\(\newcommand{\p}[1]{\frac{\partial }{\partial #1}}\) \(\newcommand{\pp}[2]{\frac{\partial #1}{\partial #2}}\) \(\newcommand{\dd}[2]{\frac{d #1}{d #2}}\) \(\newcommand{\h}{\frac{1}{2}}\) \(\newcommand{\op}[1]{\operatorname{#1}}\)
8.7.3.8. Macromolecular parameterization of growth
To enable the macromolecular parameterization of phototrophic growth, define DARWIN_MACROMOLECULAR_GROWTH in DARWIN_OPTIONS.h. Phytoplankton need to have flexible nitrogen, phosphorus and iron quotas, so also define DARWIN_ALLOW_NQUOTA, DARWIN_ALLOW_PQUOTA and DARWIN_ALLOW_FEQUOTA.
The macromolecular growth parameterization is based on [SID+26]. In order to parameterize the growth rate as a function of elemental quotas, we assume that the available carbon, nitrogen, phosphorus and iron in the organism are allocated optimally to macromolecular pools to achieve maximum growth rate, \(P_{\mathrm{C}}\), at every instant. To determine this growth rate, the macromolecular requirements are expressed as a function of growth rate. The available quota of each element then provides a constraint on the achievable growth rate which can be obtained by solving for \(P_{\mathrm{C}}\) as a function of the required quota. The realized growth rate is the smallest of these solutions, the one obtained from the most limiting quota.
The assumed macromolecular pools and fluxes between them are shown in Figure 8.24.
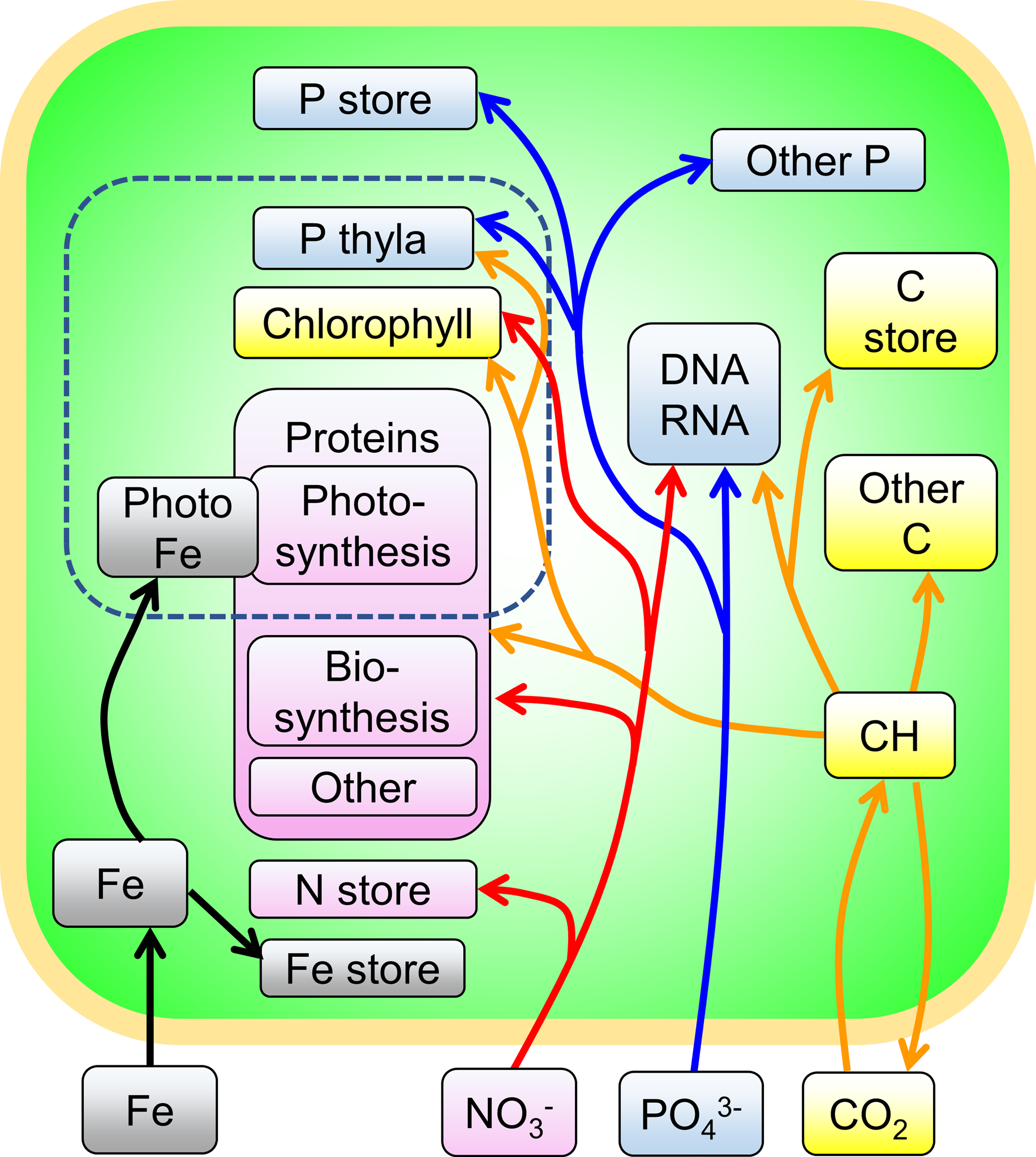
Figure 8.24 Macromolecular pools and fluxes
They imply the following elemental quota requirements:
Any quotas beyonds these essential values will be distributed between a store and an excess that is used to regulate uptake,
The molar fractions of the macromolecules are a function of light, temperature, and nutrient uptake. To find the required chlorophyll quota, \({Q_{\mathrm{C}}^{\mathrm{Chl}}}\) , consider the net change of cellular carbon quota from photosynthesis, growth and maintainance:
The chlorophyll-specific photosynthesis rate is a function of the photosynthetically active radiation, \(I\),
By assuming fixed carbon quota, the chlorophyll quota required for growth rate \(P_{\mathrm{C}}\) at given light is:
The photosynthetic protein pool is assumed to be proportional to chlorophyll,
The biosynthetic protein pool to growth rate, with a temperature dependence (only DARWIN_TEMP_VERSION 2 is supported)
Total proteins also include a base pool of ‘other’ protein,
Here, the elemental quotas of the pools are related as
RNA pools depend on protein pools and growth rate, again with a temperature dependence,
with
We find the other cell component from the following stoichiometric relations:
These cellular balances for each element, without the storage component, yield trinomial equation for growth rate, which solve numerically
By equating these to the actually available quotas, we obtain maximum growth rates for each quota limitation, \(P_{\mathrm{C}}^X\). The realized growth rate is the smallest,
8.7.3.8.1. No-growth, non-zero chlorophyll case
When one of the nutrient-limited growth-rate equations does not have a positive solution, the growth rate is zero. In this case, it is assumed that the macromolecular requirements for Chlorophyll in the first part of this section are still valid (but for \(P_{\mathrm{C}}=0\)). We can then compute the maximum amount of chlorophyll possible for each limiting nutrient,
where
are the essential quotas for zero growth. (Without growth, no iron is required).
The carbon requirements also imply a maximum chlorophyll value for the zero growth case,
where
The realized amount of chlorophyll is the one compatible with all these, and the constraint from carbon requirement,
8.7.3.8.2. No-chlorophyll case
When even the minimum quota requirements for chlorophyll are not met, the available quotas are divided evenly between the pools needed for chlorophyll,
where
Any excess (of the non-limiting element) goes to the nitrogen protein or phosphorus ‘other’ pool,
The essential quotas are then recomputed with these reduced pools.
8.7.3.8.3. Storage and uptake regulation
Nutrient storage is computed from excess quota beyond essential. Nitrogen storage is limited by a fixed maximum and the required carbon quota for the store,
For phosphorus and iron, the quotas are limited rather than the store,
Excess nitrogen quota that cannot be stored leads to a reduction in uptake via an additional multiplicative regulation term in \(U^{\mathrm{NO3}}\),
and similar for phosphorus and iron.
symbol |
trait |
param |
default |
units |
description |
|---|---|---|---|---|---|
\(E\) |
0.774 |
dimensionless |
respiratory cost of biosynthesis |
||
\(m\) |
0.393/day |
1/s |
maintenance respiration rate |
||
\(V^{\max}_{\mathrm{I}}\) |
277/day |
molC/s / (molC in Chl) |
per-chlorophyll maximum photosynthesis rate |
||
0 |
|||||
\(A_{\mathrm{I}}\) |
0.008633641 |
m2s/μmol |
coefficient characterizing the absorption cross section |
||
\(S_{\mathrm{f}}\) |
1.0 |
unitless |
enhancement of photosynthesis due to size |
||
\(A_{\mathrm{Pho}}\) |
16.0 |
molC / (molC in Chl) |
A constant of proportionality |
||
\(A_{\mathrm{Bio}}\) |
0.2711*day |
molC / (molC/s) |
constant for variable part of biosynthesis protein |
||
\(A^{\mathrm{P}}_{\mathrm{RNA}}\) |
0.00423*day |
molP / (molC/s) |
constant for variable part of RNA |
||
\(Q_{\mathrm{C}}^{\mathrm{Other}}\) |
0.0182 |
molC / molC |
constant pool of structural lipids and carbs |
||
\(Q_{\mathrm{C}}^{\mathrm{Pro\_Other}}\) |
0.24 |
molC / molC |
constant pool of essential proteins |
||
\(Q_{\mathrm{P}}^{\mathrm{Other}}\) |
6.5344E-4 |
molP / molC |
constant part of phosphorus |
||
\(Q_{\mathrm{P,min}}^{\mathrm{RNA}}\) |
2.23E-4 |
molP / molC |
minimum RNA in the cell |
||
\(Q_{\mathrm{C}}^{\mathrm{DNA}}\) |
9.41E-4 |
molC / molC |
constant part of DNA |
||
\(Q_{\mathrm{N,max}}^{\mathrm{Sto}}\) |
0.035 |
molN / molC |
maximum nitrogen storage |
||
0 |
|||||
\(Q_{\mathrm{P}}^{\mathrm{max}}\) |
0.0052 |
molP / molC |
maximum phosphorus quota |
||
0 |
|||||
\(Q_{\mathrm{Fe}}^{\mathrm{max}}\) |
2.436E-4 |
molFe / molC |
maximum iron quota |
||
0 |
|||||
\(Y^{\mathrm{C}:\mathrm{P}}_{\mathrm{Plip}}\) |
40.0 |
molC / molP |
C/P molar ratio of thylakoid membrane |
||
\(Y^{\mathrm{C}:\mathrm{N}}_{\mathrm{Pro}}\) |
5.3/1.4 |
molC / molN |
C/N molar ratio in protein |
||
\(Y^{\mathrm{N}:\mathrm{C}}_{\mathrm{Chl}}\) |
4.0/55.0 |
molN / molC |
N/C molar ratio in chlorophyll |
||
\(Y^{\mathrm{C}:\mathrm{N}}_{\mathrm{Nsto}}\) |
2.0 |
molC / molN |
C/N molar ratio of cyanophycin |
||
\(Y^{\mathrm{P}:\mathrm{N}}_{\mathrm{RNA}}\) |
1/3.75 |
molP / molN |
P/N molar ratio of RNA |
||
\(Y^{\mathrm{C}:\mathrm{N}}_{\mathrm{DNA}}\) |
9.75/3.75 |
molC / molN |
C/N molar ratio of DNA |
||
\(Y^{\mathrm{C}:\mathrm{N}}_{\mathrm{RNA}}\) |
9.50/3.75 |
molC / molN |
C/N molar ratio of RNA |
||
\(Y^{\mathrm{P}}_{\mathrm{Thy}}\) |
0.028163 |
molP / (molC in Chl) |
phosphorus in thylakoid membrane to chlorophyll |
||
\(Y^{\mathrm{Fe}:\mathrm{N}}_{\mathrm{Pho}}\) |
0.00163 |
molFe / molN |
Fe/N ratio in photosystem iron |
symbol |
trait |
units |
description |
|---|---|---|---|
\(Q_{\mathrm{N}}^{\mathrm{Pro\_Other}}\) |
molN / molC |
cellular nitrogen in essential proteins |
|
\(Q_{\mathrm{N,min}}^{\mathrm{RNA}}\) |
molN / molC |
constant part of RNA in nitrogen |
|
\(Q_{\mathrm{C,min}}^{\mathrm{RNA}}\) |
molN / molC |
constant part of RNA in carbon |
|
\(Q_{\mathrm{N}}^{\mathrm{DNA}}\) |
molN / molC |
DNA in nitrogen |
|
\(Q_{\mathrm{P}}^{\mathrm{DNA}}\) |
molP / molC |
DNA in phosphorous |
|
\(A_{\mathrm{Thy}}\) |
molC / (molC in chl) |
ratio of carbon in thylakoid membrane to chlorophyll |
|
\(A^{\mathrm{N}}_{\mathrm{RNA}}\) |
s molN / molN |
constant for variable part of RNA |
|
\(V^{\min}_{\mathrm{I}}\) |
molC/s / (molC in Chl) |
minimum photosynthesis rate |
|
\(Q_{\mathrm{C,max}}^{\mathrm{Chl}}\) |
molC / molC |
maximum chlorophyll concentration at minimum light |
|
\(Q_{\mathrm{N}}^{\text{no-Chl}}\) |
molN / molC |
minimum QN at zero growth rate |
|
\(Q_{\mathrm{P}}^{\text{no-Chl}}\) |
molP / molC |
minimum QP at zero growth rate |
|
\(Q_{\mathrm{Fe}}^{\text{no-Chl}}\) |
molFe / molC |
minimum QFe at zero growth rate |
|
\(Q_{\mathrm{C}}^{\mathrm{const}}\) |
molC / molC |
constant portion of the cell |
param |
default |
units |
description |
|---|---|---|---|
–8420 |
K |
slope for pseudo-Arrhenius for macromolecular (TEMP_VERSION 2) |